「糖鎖や核酸などの非タンパク質抗原への獲得免疫応答」をメインテーマとして独創性の高い研究をおこなっています。また、自己免疫疾患の新規治療法の開発や、がん免疫やアレルギーを制御するワクチンの開発を行なっています。
目次
感染免疫および自己免疫疾患における非タンパク質抗原へのB細胞の応答の重要性
B細胞は核酸や多糖、糖脂質などの非タンパク質抗原も認識し、特異的な抗体を産生します。一方、T細胞は、一部の糖脂質を除き、非タンパク質抗原を認識することはできません。このため、非タンパク質抗原への獲得免疫応答はもっぱらB細胞のみによって行われます。非タンパク質抗原への抗体は、肺炎球菌など莢膜をもつ細菌への感染防御で中心的な役割を果たす一方で、全身性エリテマトーデス(SLE)やギラン・バレー症候群などの自己免疫疾患では、非タンパク質自己抗原への自己抗体産生が疾患発症で重要な役割を果たします。
非タンパク質抗原へのB細胞応答の仕組み
教科書では、B細胞が抗体を産生するには同じ抗原を認識する活性化T細胞による補助が必要です。T細胞は、一部の糖脂質を除き、非タンパク質抗原を認識することはできず、このため、一般的にT細胞は非タンパク質抗原を認識したB細胞の活性化を補助することができません。非タンパク質抗原には内因性のB細胞活性化シグナルが存在し、この内因性シグナルによってT細胞からの補助がなくても抗原に反応したB細胞が活性化し、抗体産生を行うと考えられます。核酸はTLR7などの核酸センサーにより認識され、活性化シグナル伝達を誘導する。しかし、多糖や糖脂質などの糖鎖関連抗原の内因性活性化シグナルがどのようなものかはまだ未解明です。私たちの研究グループでは、現在、多糖がT細胞の補助なしでB細胞を活性化する仕組みの解明を行なっています。
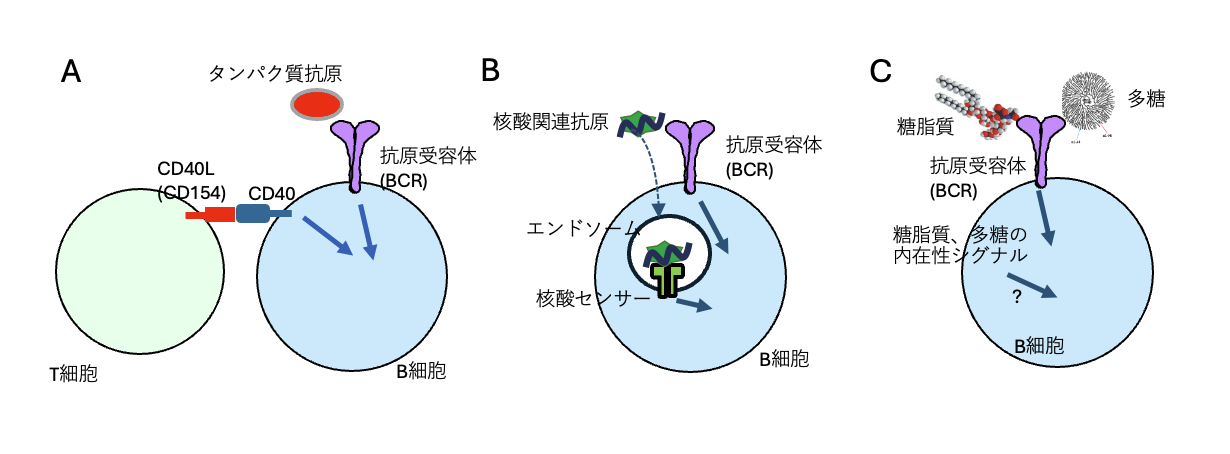
図1.異なる抗原による異なるB細胞の活性化の仕組み
(A) タンパク質抗原を認識したB細胞のT細胞依存的な活性化 タンパク質抗原に反応したB細胞は抗原受容体(BCR)を介するシグナル伝達を誘導するとともに、同じ抗原に反応するT細胞と相互作用する。この相互作用の過程で、T細胞のCD40L(CD154)がB細胞のCD40に結合すし、CD40を介する活性化シグナル伝達が誘導される。この活性化シグナルとBCRを介するシグナル伝達によりB細胞は活性化する。(B)核酸関連抗原を認識したB細胞のT細胞非依存的な活性化 核酸や核酸を含む複合抗原(核酸関連抗原)をB細胞が認識すると、BCRを介するシグナル伝達が誘導される。さらに、BCRを介するエンドサイトーシスにより抗原が細胞内に取り込まれ、エンドソームでTLR7, TLR9などの核酸センサーに反応し、核酸センサーからの活性化シグナル伝達を誘導する。このシグナル伝達とBCRを介するシグナル伝達によりB細胞は活性化する。(C)糖脂質や多糖によるT細胞非依存的なB細胞の活性化。糖脂質や多糖にはこれらの抗原に内在するB細胞を活性化させるシグナルが存在し、これらの抗原を認識したB細胞を活性化するが、内在性シグナルの本体は不明である。
非タンパク質自己抗原への自己免疫を防止する仕組みの解明
私たちの研究グループでは、B細胞が発現する抑制性受容体の研究を行なってきました。CD72はもっぱらB細胞に発現する抑制性受容体で、ヒトのSLEやマウスのSLE様の自己免疫疾患と遺伝的に関連し、さらに、海外の研究グループや私たちの研究グループではCD72を欠損するマウスを樹立し、このマウスでSLEを自然発症することを明らかにし、CD72がSLEの発症を抑制することを明らかにしました。SLEでは、種々の核酸を含む複合抗原への自己抗体が産生され、疾患発症に関わります。この中でもとりわけ、Sm/RNP複合体やリボソームへの自己抗体はSLE以外のリウマチ疾患では認められませんので、SLEに特徴的な自己抗体と言えます。私たちは、CD72がSm/RNPやリボソームに結合し、B細胞の活性化を抑制することで、これらの自己抗原に反応したB細胞の自己寛容を誘導することを明らかにしました。
ギラン・バレー症候群では、ニューロンに発現するガングリオシドと呼ばれる一群の糖脂質に対する自己抗体が産生され、末梢神経障害がおこります。私たちは、B細胞に発現する抑制性受容体Siglec-10が種々のガングリオシドを認識することを明らかにしました。また、稀なSiglec-10の機能喪失(loss-of-function)変異を発見し、この変異がギラン・バレー症候群と強く関連することを明らかにしました。この知見から、Siglec-10がガングリオシドを認識し、B細胞のガングリオシドへの自己抗体産生を抑制することで、ギラン・バレー症候群の発症を抑制することが示唆されます。
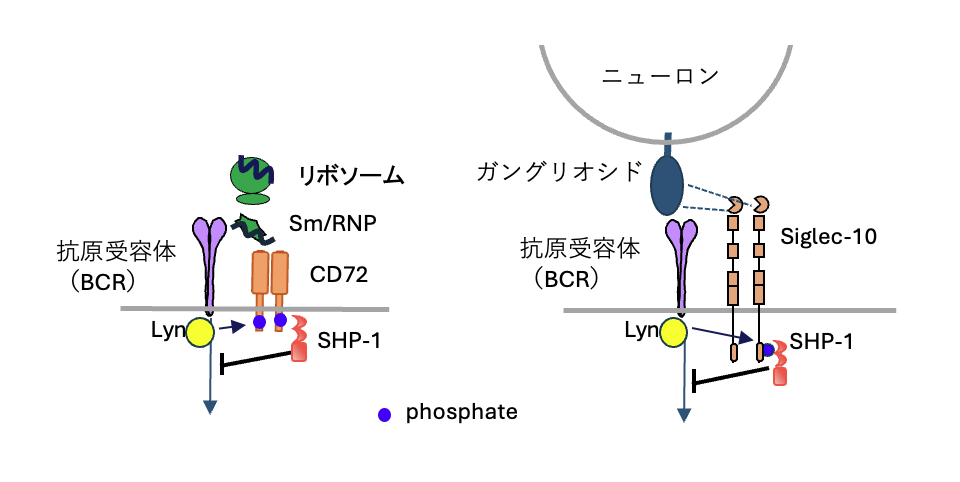
図2 抑制性受容体による核酸関連自己抗原および糖脂質自己抗原へのB細胞トレランスの誘導
B細胞はCD72やSiglec-10などの抑制性受容体を発現する。CD72は、核酸を含む自己抗原で全身性エリテマトーデス(SLE)での自己抗体の標的となるSm/RNPやリボソームを認識し、これらの自己抗原を認識したB細胞の活性化を抑制する。また、Siglec-Gは、ギラン・バレー症候群で産生される自己抗体の標的分子であるガングリオシドを認識し、ガングリオシドを認識したB細胞の活性化を抑制する。これらの抑制性受容体によるB細胞の活性化抑制には、抗原を認識した抗原受容体(BCR)により活性化したチロシンキナーゼによる、抑制性受容体の細胞内領域のリン酸化、このリン酸化部位によるチロシンホスファターゼSHP-1の活性化とSHP-1による抗原受容体を介するシグナル伝達の抑制が関わる。その結果、これらの非タンパク質自己抗原へのB細胞トレランスが誘導され、自己抗体産生が抑制される。
B細胞の自己寛容はT細胞の自己寛容により制御され、B細胞が自己抗原を認識しても、T細胞からの補助がないために、自己反応性B細胞は通常は活性化しません。しかし、B細胞が自己の非タンパク質抗原や非タンパク質抗原を含む複合抗原を認識した際に、T細胞による補助がなくても非タンパク質抗原が持つ内因性活性化シグナルによりB細胞を活性化し、自己抗体産生がおこります。そこで、CD72やSiglec-10が自己非タンパク質自己抗原を認識し、非タンパク質自己抗原へのB細胞応答を抑制することで、T細胞自己寛容によって制御できないB細胞の自己寛容を誘導するものと考えられます。
SLEでは、Sm/RNPやリボソーム以外にもヌクレオソームなど種々の自己抗原への自己抗体産生がおこり、CD72欠損マウスでも同様に種々の自己抗原への抗体産生がおこります。私たちの研究グループでは、CD72がどのようにして種々の自己抗原への自己抗体産生を抑制するのか、現在その仕組みの解明を行なっています。また、この仕組みを増強することでSLEを治療する新規医薬品の開発を行なっています。
自己免疫疾患の新規治療薬の開発
一部のB細胞はIL-10などの抑制性サイトカインを分泌することで免疫応答を抑制する。このようなB細胞は制御性B細胞(Breg細胞)と呼ばれ、種々の自己免疫疾患でBreg細胞の数や機能が減弱していることが示されている。また、Bregが移植片拒絶を抑制することも明らかになってきている。IL-6やTLRのリガンドがBreg細胞を増強することが知られているが、これらは炎症を増悪するため、自己免疫疾患の治療に用いることはできない。私たちは、Breg細胞を増強するが炎症を誘導しない標的分子を発見し、この分子への抗体によりマウスの自己免疫疾患の発症抑制や移植片拒絶を抑制することを明らかにしました。現在、このような抗体を用いた自己免疫疾患の新規治療薬の開発を行なっている。
また、私たちの研究グループでは、CD72の機能増強によるSLEの治療薬開発を行なっています。
抗体医薬に代わる抗体産生治療ワクチンの開発
近年の画期的な治療薬の多くは、抗体医薬です。これは、低分子化合物が標的とできないような分子も抗体医薬によって制御できるからです。逆に、低分子化合物で抗体医薬の代替を行なうことは極めて困難です。しかし、抗体医薬は非常に高価になるため、何らかの代替法が求められています。標的分子の一部をワクチンとして免疫することで、患者さん自身が標的分子に対する抗体を作り、抗体医薬と同等の効果を得ることが可能と考えられています。私たちは、このような抗体医薬に代わる抗体産生治療ワクチン開発の基盤技術の開発を行っています。この技術により、抗体医薬の導入というパラダイムシフトに匹敵するような、疾患治療のパラダイムシフトが起こると期待されます。
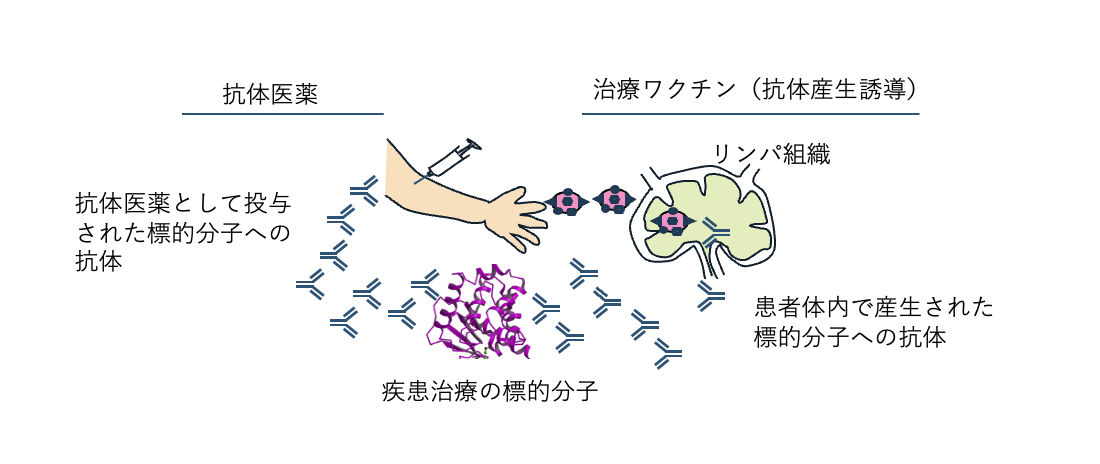
図3. 抗体医薬に代わる抗体産生誘導治療ワクチン
抗体医薬が患者に投与されると、その成分である抗体が標的分子に特異的に結合し、治療効果が発揮される。抗体産生誘導ワクチンは、治療標的分子の一部を含み、患者に投与さえると、標的分子への抗体産生を誘導する。この抗体が抗体医薬と同様の効果を発揮する。